
VI. ВЕЛИКИЕ СТОХАСТИЧЕСКИЕ ПРОЦЕССЫ
6. ГОМОЛОГИЯ
Здесь я расстаюсь с проблемами индивидуальной генетики, соматического изменения и обучения, и непосредственными путями эволюции, чтобы рассмотреть результаты эволюции в более крупном масштабе. Я займусь теперь вопросом о том, что можно сказать о соответствующих процессах с точки зрения более широкой картины филогенеза.
Сравнительная анатомия имеет долгую историю. В течение по меньшей мере шестидесяти лет, от опубликования «Происхождения видов» до 1920-х годов, сравнительная анатомия была сосредоточена на взаимосвязях, не рассматривая процессов. Возможность построить филогенетические древа, как полагали, свидетельствовала в пользу теории Дарвина. Ископаемые данные были неизбежно весьма неполны, и в отсутствие прямых свидетельств о происхождении, анатомы проявляли ненасытный интерес к определенному классу сходства, именуемого гомологией. Гомология «доказывала» взаимосвязь, а взаимосвязь и была эволюцией.
Конечно, люди замечали формальное сходство живых существ по крайней мере с начала эволюции языка, соединявшего мою «руку» с вашей «рукой» и мою «голову» с «головой» рыбы. Но лишь гораздо позже возникло понимание необходимости объяснить такие формальные сходства. Даже в наше время большинство людей не удивляется сходству между двумя руками и не видит в этом никакой проблемы. Они не чувствуют и не усматривают никакой надобности в теории эволюции. Для мыслящих людей древности, и даже людей Возрождения формальное подобие живых существ было иллюстрацией связности Великой Цепи Бытия, и эти связи были для них логическими, а не генеалогическими связями.
Так или иначе, скачкообразное заключение от формального сходства к взаимосвязи скрывало ряд скачкообразных гипотез.
ПризнAем формальное сходство в тысячах случаев — между человеком и лошадью, омаром и крабом — и допустим, что в этих случаях формальное сходство — не только свидетельство, но попросту результат эволюционного отношения. Тогда мы можем рассмотреть вопрос, проливает ли природа сходства в этих случаях свет на процесс эволюции.
Спросим себя: что говорят нам гомологии о процессе эволюции? При сравнении нашего описания омара с нашим описанием краба мы обнаруживаем, что некоторые компоненты описаний остаются неизменными, а другие отличаются друг от друга. Поэтому первый шаг несомненно должен принять форму различения между разными видами изменения. Некоторые изменения будут подчеркнуты как более вероятные и более легкие; другие окажутся более трудными, и потому менее вероятными. В таких условиях медленно меняющиеся переменные будут отставать и могут стать ядром тех гомологий, на которых можно основывать более широкие гипотезы таксономии.
Но эта первая классификация изменений на быстрые и медленные и сама потребует объяснения. ЧтO можно прибавить к нашему описанию эволюционного процесса, что могло бы, возможно, предсказать, какие переменные будут в самом деле медленно меняющимися и станут, тем самым, базисом гомологии?
Насколько мне известно, единственные начала такой классификации неявно содержатся в так называемой теории повторения.
Зародышем доктрины повторения было высказывание одного из пионеров эмбриологии, немца Карла Эрнста фон Бера в 1828 году о «законе соответствующих стадий». Он доказывал свой закон с помощью приема сравнения эмбрионов позвоночных разных непомеченных видов.
Я никак не в состоянии определить, к какому классу они принадлежат. Это могут быть ящерицы, или небольшие птицы, или очень молодые млекопитающие — столь полно сходство в способе формирования головы и туловища этих животных. Конечности еще отсутствуют, но если бы даже они существовали, на ранней стадии развития мы ничего бы не узнали, поскольку все возникает из одной и той же основной формы. [Encyclopedia Britannica, статья «Baer, Karl Ernst von (1792–1876)».]
Концепция «соответствующих стадий» фон Бера была позже разработана Эрнстом Геккелем, современником Дарвина, в теорию повторения и в часто оспаривавшееся утверждение, что «онтогенез повторяет филогенез». С тех пор были предложены весьма разнообразные формулировки этого положения. Самое осторожное из них — это, пожалуй, утверждение, что личинки или эмбрионы данного вида обычно более напоминают личинки или эмбрионы родственного вида, чем взрослые особи напоминают взрослые особи родственного вида. Но даже эта очень осторожная формулировка ставится под сомнение заметными исключениями. [Например, среди червообразных морских животных прежнего класса Enteropneusta различные виды, которые относили к одному роду Balanoglossus, имеют совершенно разную эмбриологию. B.Kovalevskii имеет похожих на головастиков личинок, с жаберными щелями и хордой, между тем как другие виды имеют личинок, напоминающих личинки иглокожих.]
Но несмотря на эти исключения я склонен думать, что обобщение фон Бера составляет важный ключ к пониманию процесса эволюции. Это обобщение — верно оно или неверно — ставит важные вопросы о выживании не организмов, а признаков: Существует ли общий наибольший знаменатель тех переменных, которые становятся устойчивыми, и поэтому использовались зоологами в их поиске гомологии? Закон соответствующих стадий имеет то преимущество перед более поздними формулировками, что не претендует на филогенетические древеса, и даже краткая цитата, приведенная выше, содержит некоторые особенности, которых не заметит филогенетический изыскатель. Верно ли, что эмбриональные переменные более стойки, чем переменные взрослых?
Фон Бер сосредоточивается на высших позвоночных — ящерицах, птицах и млекопитающих — животных, эмбриология которых поддерживается и защищается либо наполненным пищей яйцом, либо материнской утробой. Но доказательства фон Бера просто не действовали бы, скажем, для личинок насекомых. Любой энтомолог, взглянув на не снабженную надписями выставку личинок жуков, может сразу сказать, к какому семейству принадлежит каждая личинка. Разнообразие личинок столь же удивительно, как и разнообразие взрослых особей.
Закон соответствующих стадий, по-видимому, верен не только для целых эмбрионов позвоночных, но также и для последовательности конечностей на самых ранних стадиях их развития. Так называемая серийная гомология разделяет с филогенетической гомологией то свойство, что, в общем, сходства предшествуют различиям. Вполне развитая клешня омара заметно отличается от ходячих конечностей на четырех других сегментах груди, но все конечности груди на их ранней стадии выглядят похоже.
Возможно, именно это и есть граница, до которой следует доводить обобщение фон Бера: можно утверждать, что, вообще говоря, сходство старше различия (и в филогенетическом, и в онтогенетическом смысле). Некоторым биологам это может показаться трюизмом, как если сказать, что в каждой ветвящейся системе две точки, близкие к точке ветвления, будут более сходны между собой, чем другие две точки, далекие от нее. Но этот кажущийся трюизм не будет справедлив для элементов периодической системы, и не обязательно будет справедлив в биологическом мире, произведенном намеренным творением. В действительности наш трюизм свидетельствует в пользу гипотезы, что организмы в самом деле связаны между собой, как точки или положения на ветвящемся дереве.
Обобщение, по которому сходство старше различия, — все еще весьма неполное объяснение наличия гомологии в тысячах примеров, во всем биологическом мире. Говоря, что сходства старше различий, мы только повторяем вопрос: «Почему некоторые признаки становятся основой гомологии?». Вопрос этот почти не меняется, принимая следующий вид: Почему некоторые признаки становятся старшими, сохраняясь дольше, и тем самым становятся основой гомологии?
Мы сталкиваемся здесь с проблемой выживания — не выживания видов или разновидностей, борющихся во враждебном мире других организмов, а с более тонкой проблемой выживания признаков (предметов описания), которые должны выжить и во внешнем окружении, и во внутреннем мире других признаков, во всей совокупности репродукции, эмбриологии и анатомии данного организма.
Почему же некоторые части описания, входящие в составленную ученым сложную систему описания всего организма, остаются в силе дольше других частей (в течение ряда поколений)? И есть ли совпадение, наложение или синонимия между частями и кусками описания и частями и кусками набора инструкций, определяющего онтогенез?
Если бы слон имел строение зубов и другие формальные признаки семейства Muridae [Семейство грызунов, включающее мышей и крыс. — Прим. перев.], он был бы мышью, несмотря на свой размер. В самом деле, даман, размером с кошку, близок к тому, чтобы быть гиппопотамом, а лев очень близок к тому, чтобы быть кошкой. Размер сам по себе, по-видимому, несуществен. ЧтO важно — это форма. Но не так легко определить, что понимается под «формой» или «паттерном» в этом контексте.
Мы ищем критерии, позволяющие распознать признаки, которые, как можно думать, дольше остаются справедливыми в сумятице эволюционного процесса. Здесь выделяются два признака — два традиционных способа подразделения в обширном поле «различий». Существует дихотомия между паттерном и количеством и дихотомия между непрерывностью и разрывностью. Соединяются ли контрастирующие организмы непрерывным рядом шагов, или же отделены резким переходом? Затруднительно (хотя и возможно) представить себе постепенный переход между паттернами, а потому эти две дихотомии, по-видимому, перекрываются. Во всяком случае, можно ожидать, что теоретики, предпочитающие ссылаться на паттерны, будут также предпочитать теории, ссылающиеся на разрывность. (Но, конечно, подобные предпочтения, зависящие лишь от умонастроения отдельного ученого или от модного мнения, заслуживают порицания).
Я полагаю, что самые убедительные открытия в этом вопросе — это изящные доказательства зоолога Д’Арси Вентворта Томпсона, предложенные в начале двадцатого века. Как он показал, во многих случаях — пожалуй, во всех рассмотренных им случаях — две контрастирующих, но родственных формы животных имеют следующее общее свойство: если одна из этих форм изображена (скажем, в профиль) в декартовых ортогональных координатах (например, на разграфленной бумаге), то, при надлежащем наклоне или искажении, те же координаты подойдут и к другой форме. Все точки профиля второй формы совпадут с одноименными точками в искаженных координатах (см. рис. 9).
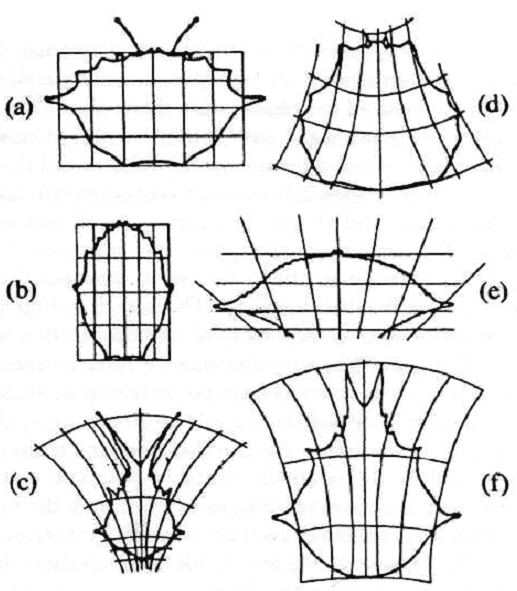
Рис. 9
Примечательная особенность открытий Д’Арси Томпсона состоит в том, что в каждом случае искажение неожиданно просто, причем оно последовательным образом сохраняется в изображении всего животного. Наклон координат таков, что его можно описать некоторым простым математическим преобразованием.
Эта простота и последовательность должны несомненно означать, что различия между фенотипами, выявляемые методом Д’Арси Томпсона, представляют лишь немногие различия в генотипе (т. е. в небольшом числе генов).
Более того, последовательность искажения, распространяющаяся на все тело животного, по-видимому, указывает, что относящиеся к этому гены плейотропны (т. е. влияют на многие, может быть, на все части фенотипа), притом такими способами, которые в этом смысле гармонизированы по всему телу.
Дальнейшая интерпретация этих открытий не совсем проста, и сам Д’Арси Томпсон в этом не очень помогает. Его восхищает тот факт, что математика оказалась способной описать некоторые виды изменений.
Рисунок заимствован из книги Д’Арси Томпсона «О росте и форме» (D’Arcy Tompson, On Growth and Form), стр. 294. Cambridge University Press, 1961.
В этой связи любопытно отметить нынешний спор между сторонниками «синтетической» теории эволюции (т. е. нынешнего ортодоксального дарвинизма) и их противниками, «типологами». Например, Эрнст Майр высмеивает слепоту типологов: «Как показывает история, типолог нисколько не оценивает, и не способен оценить естественный отбор». [См. Ernst Mayr, Populations, Species and Evolution (Cambridge, Mass.: Harvard University Press, 1963), p. 107.] К сожалению, он не ссылается на источники своей типологии, применяемой им к его коллегам. Может быть, он слишком скромен, чтобы претендовать на эту честь? Или же, в этом случае, похожие узнают друг друга?
Но разве не все мы в душе типологи?
Во всяком случае, несомненно, есть много способов рассматривать формы животных. И поскольку мы принялись, в духе Платона, за исследование параллелизма между творческим мышлением и обширным разумным (mental) процессом, именуемым биологической эволюцией, то в каждом случае стOит спросить: Не представлен ли этот способ рассмотрения явлений, не параллелен ли он в некотором роде чему-то в системе организации самих явлений? Нет ли в каких-то генетических сообщениях и устойчивых признаках, определяющих фенотип, чего-то вроде синтаксиса (за неимением лучшего слова), отделяющего «типологическое» мышление от «синтетического»? Можем ли мы распознать среди множества сообщений, создающих и определяющих формы животных, некоторые более типологические и более синтетические?
Если поставить вопрос таким образом, то представляется, что Томпсон [Здесь мы исправили несомненную из контекста описку автора, заменившего фамилию Томпсона на Майра. — Прим. перев.] глубоко прав, предлагая свою типологию. Старые рисунки Д’Арси Томпсона именно разделяют два вида коммуникаций внутри самого организма. Рисунки показывают, что у животных есть два вида признаков: (а) У них относительно устойчивые квази-топологические [В математической дисциплине топологии изучаются качественные признаки фигур, не зависящие от их размеров и не меняющиеся при их непрерывных преобразованиях. — Прим. перев.] паттерны, которые понятным образом побудили ученых постулировать резкую разрывность в процессе эволюции. Эти признаки остаются неизменными под действием (б) относительно неустойчивых количественных свойств, видимых при сравнении смежных изображений.
Если начертить координатные линии, соответствующие квази-топологическим свойствам, то оказывается, что изменения менее устойчивых свойств представляются искажением координат.
В смысле рассматриваемого вопроса о гомологии, складывается впечатление, что в самом деле существуют разные виды свойств, и что филогенетическая гомология несомненно зависит от более устойчивых, квази-топологических паттернов.










