
Часть 2. Методологический анализ психологических теорий
2.2. От Павлова к Бернштейну[39]
Н.А. Бернштейн: физиология активности
В наши задачи не входит сколько-нибудь подробный анализ концепции Н.А. Бернштейна. Думается, она вообще должна еще подождать своего историка — слишком мало мы отошли от того грандиозного события в развитии физиологического (а косвенно — и психологического) мышления, которое являет собой физиология активности. Тем не менее уже сейчас можно выделить те главные открытия, которые сделал Н.А. Бернштейн.
1. Факт сложности движения
Краеугольным камнем теоретических построений Н.А. Бернштейна является описанный им факт неоднозначной связи между эфферентным импульсом и результирующим движением. Чтобы проследить за логикой мысли Н.А. Бернштейна47, нужно начать с элементарного вопроса: что непосредственно обеспечивает судьбу особи в процессе приспособления к реальным обстоятельствам жизни? Ответ очевиден: эффекторные функции. Рецепторика же представляет собой подсобную, обслуживающую функцию. «Нигде в филогенезе созерцание мира не фигурировало как самоцель, как нечто самодовлеющее» (Бернштейн, 1947,с.9).
47 Увы, приходится браться за неблагодарное дело: пересказывать логически безупречные и стилистически поэтичные работы Н.А Бернштейна (не зря в свое время автор учился на филологическом) — значит неизбежно огрублять их.
При этом животное, обитающее и действующее в среде, своим внешним поведением вынуждено «говорить на языке этой среды». Это необходимое условие его приспособленности к внешнему миру. Кроту необходимо прорыть ход в почве, обладающей совершенно конкретными физическими характеристиками, рыси — приземлиться при прыжке именно в ту точку и в то время, когда там находится жертва, а той, чтобы выжить, нужно суметь убежать. Иначе говоря, в процессе приспособления животное постоянно решает «двигательные задачи». Двигательная задача — это определяющееся совокупной ситуацией изменение предметной действительности, которое необходимо осуществить животному в данный момент. Для того чтобы решить ее, естественно, нужно осуществить ряд движений.
Рассмотрим, как связаны между собой эти движения и эфферентная импульсация, которая их вызывает. Для упрощения изложения этой проблемы введем понятие «парциального» движения (хотя сам Н.А. Бернштейн им не пользуется) по аналогии с понятием парциального давления газа в физике. Парциальным будем называть такое гипотетическое движение, которое бы произошло, если бы во время движения тело оказалось бы вне поля действия каких-либо внешних сил. Иначе: парциальное движение мы получили бы, если бы из совокупности всех сил, определивших реальное движение, вычли все вектора внешних сил. Введением этого понятия фиксируется тот тривиальный, казалось бы, факт, что реальное движение тела зависит не только от сил внутренних, но и от внешних по отношению к организму сил (тяготения, сопротивления предметов и противников и т. д.).
Что представляют собой эти последние? Определяется ли парциальное движение только сокращением мышц? Стоит лишь попристальней взглянуть на тело как на механическую систему, чтобы убедиться, что это не так. То, каково будет парциальное движение, зависит и от положения, в котором находилось тело в момент сокращения мышц, и от тех относительных ускорений, с которыми двигались его отдельные звенья. То есть парциальное движение определяется сокращением мышц и инерцией тела. Но более того: оказывается, что само сокращение мышцы зависит не только от центрального импульса, но также от функционального состояния мышцы и от ее наличной длины (Бернштейн, 1966, с. 21).
(Напомним, что между напряжением мышцы и ее наличной длиной существует механическая обратная связь: изменение напряжения мышцы изменяет распределение сил в сочленении, следовательно, и взаимораспределение и взаиморасположение звеньев сочленения, а значит, и длину мышцы.)
Таким образом, писал Н.А. Бернштейн, преодолевается «старое привычное представление, implicite принятое и до сих пор сохранившееся у многих физиологов и клиницистов, согласно которому скелетное звено вполне покорно центральному импульсу и однозначно повинуется ему. По этому представлению центральный импульс "а" всегда вызывает движение "А", а импульс "в" — движение "В", из чего далее следует, что легко строится представление о двигательной зоне коры как о распределительном пункте с пусковыми кнопками» (Бернштейн, 1966,с. 43).
Этой абстракции «простого движения» Н.А. Бернштейн противополагает представление, которое можно было бы назватьмоделью сложного движения. Попробуем изобразить ее в виде схемы 1.
Эта схема упрощает строгие математические и инженерные выкладки Н.А. Бернштейна, но и в таком виде она фиксирует главное — факт, что между центральной эфферентной импульсацией и реальным движением животного существует связь сложная, неоднозначная и принципиально невычислимая до деятельного столкновения животного с конкретной предметной ситуацией. Это и есть факт сложности движения, как мы его назвали. Тем самым была преодолена одна из основных догм традиционных рефлексологических и бихевиористских теорий, полагавших, что любое движение животного содержится в потенциальной форме в организме, высвобождаясь в неизменном виде при наличии определенной, раз и навсегда заданной эфферентной импульсации.
К методологическим следствиям этого открытия у нас еще будет повод обратиться, но у медали факта сложности движения есть и конкретно-теоретическая обратная сторона — сложность восприятия.
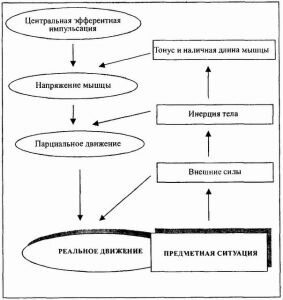
Схема 1. Модель сложного движения











