
Глава 13 СОН И СНОВИДЕНИЯ
1. АКТИВНОЕ НАСТУПЛЕНИЕ СНА ИЛИ ЛИШЕНИЕ БОДРСТВОВАНИЯ?
Уже в самых ранних исследованиях механизмов сна чётко намечаются две основные точки зрения на эту проблему. Первая — сон возникает в результате активного процесса, возбуждения определённых структур («центров сна»), которое вызывает общее снижение функций организма (активные теории сна). Вторая — это пассивные теории сна, или теории деафферентации, согласно которым сон наступает пассивно в результате прекращения действия каких-то факторов, необходимых для поддержания бодрствования. Различия между этими направлениями удачно определил Н. Клейтман, писавший, что «заснуть» и «не суметь остаться бодрствующим», — это не одно и то же, так как первое предполагает активное действие, а второе — пассивное устранение активного состояния.
В первых неврогенных теориях сна, возникших в конце прошлого и начале нынешнего столетия в результате клинических наблюдений после эпидемии летаргического энцефалита, отражаются эти две противоположные тенденции. Так, Л. Маутнер пришёл к выводу, что причиной наступления сна при этом заболевании была опухоль в области среднего мозга, вызывавшая сдавление афферентных путей и прекращение потока сенсорных импульсов в высшие отделы мозга. Фон Экономо же считал, что сон наступает в результате торможения таламуса и коры больших полушарий «центром сна». Исходя из противоположного характера ведущих симптомов заболевания (сонливости или бессонницы) он полагал, что центр сна состоит, по крайней мере, из двух частей — каудальной части, являющейся как бы центром бодрствования, и ростральной, действующей как центр сна.
Первыми экспериментальными исследованиями, свидетельствующими о существовании центра сна, явились работы В. Гесса. Показав, что слабое электрическое раздражение чётко ограниченной области промежуточного мозга у подопытных кошек вызывало сон со всеми подготовительными фазами (потягивание кошки, умывание, принятие характерной позы), В. Гесс высказал предположение, что существует центр, возбуждение которого обеспечивает наступление естественного сна. В дальнейшем опыты В. Гесса были подтверждены многочисленными исследователями, которые вызывали наступление сна у подопытных животных при помощи электрического и химического раздражения гипоталамуса и прилежащих структур, и теория о центре сна получила значительное признание.
Однако против подобного локализационистского объяснения механизма возникновения сна решительно выступал И.П. Павлов. Он рассматривал сон как результат торможения коры больших полушарий; при этом его теория сна не исключала участия и подкорковых структур в возникновении сна. В качестве основных этапов в дальнейшем развитии «пассивных» теорий можно отметить работы Н. Клейтмана, который пришёл к выводу о том, что сон — это «легко обратимая пассивность высших функциональных центров коры», которая наступает в результате уменьшения потока афферентной импульсации. Эксперименты Ф. Бремера с поперечными перерезками на разных уровнях ствола мозга кошки, показавшие, что межколликулярная перерезка приводит к картине сна на ЭЭГ (спящий «cerveau isole»), тогда как перерезка на границе головного и спинного мозга не изменяет картину бодрствования на ЭЭГ (бодрствующий «encephale isole»), подтверждали представление о сне как результате деафферентации коры больших полушарий. В результате опытов с разрушением различных областей промежуточного мозга у обезьян, приводивших к наступлению сна, С. Рэнсон пришёл к выводу о том, что гипоталамус является центром «интеграции эмоционального выражения» и сон наступает в результате периодического снижения активности этого центра бодрствования.
Открытие Дж. Моруцци и X. Мэгуном в 1949 г. восходящего активирующего влияния неспецифической ретикулярной системы (ВРАС) значительно усилило позиции пассивных теорий сна. Поддержание бодрствующего состояния объяснялось теперь тоническим влиянием ВРАС. Дальнейшие исследования привели к открытию и других активирующих систем — диффузной и специфической таламической систем и активирующих структур заднего гипоталамуса (см. гл. 8).
Впоследствии в целом ряде работ было показано, что раздражение ретикулярных структур ствола мозга может вызывать не только пробуждение, но и сон. Так, низкочастотное раздражение почти всех исследуемых пунктов ретикулярной формации (область среднего мозга, моста и продолговатого мозга) у кошек вызывало наступление сна, что позволило Дж. Росси и А. Цанкетти предположить существование в стволе мозга двух антагонистических механизмов — десинхронизирующего (пробуждающего) и синхронизирующего (вызывающего сон), — поэтому сон не пассивное состояние отсутствия бодрствования, а результат активного торможения деятельности мозга. Моруцци также привёл экспериментальные доказательства того, что в нижних отделах ствола мозга имеются синхронизирующие структуры, действующие как антагонисты ВРАС. К синхронизирующим структурам, раздражение которых может вызвать наступление сна у животного, позже были отнесены и хвостатое ядро, и неспецифическая таламическая система, и структуры базальной части переднего мозга, в частности, преоптическая область (подробнее см. обзоры [Вейн и др., 1971; Шевченко, 1971; Блум и др., 1988]).
Одна из попыток создания единой теории сна была предпринята П.К. Анохиным [1945, 1968]. Состояние сна он представлял как результат проявления целостной деятельности организма, строго координирующей корковые и подкорковые структуры в единую функциональную систему. В своей гипотезе П.К. Анохин исходил из того, что гипоталамические «центры сна» находятся под тоническим угнетающим влиянием со стороны коры больших полушарий. Именно поэтому при ослаблении этого влияния вследствие снижения рабочего тонуса корковых клеток («активный сон» по Павлову) гипоталамические структуры как бы «высвобождаются» и определяют всю ту сложную картину перераспределения вегетативных компонентов, которая характерна для состояния сна. При этом гипоталамические центры оказывают угнетающее влияние на восходящую активирующую систему, прекращая доступ в кору всего комплекса активирующих воздействий (и наступает «пассивный сон» по Павлову). Эти взаимодействия представляются циклическими, поэтому состояние сна может быть вызвано искусственно (или в результате патологического процесса) воздействием на любую часть этого цикла (рис. 13.1).
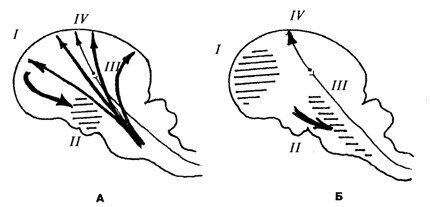
Рис. 13.1. Схема взаимодействия «центров сна» и «пробуждающих» структур при бодрствовании и наступлении сна (по П.К. Анохину)
А. Бодрствование. Корковые влияния (I) тормозят «центры сна» (II) и восходящие активирующие влияния ретикулярных структур (III) и возбуждения, идущие по лемнисковым путям (IV), свободно достигают коры. Б. Сон. Заторможенные отделы коры (I) перестают оказывать сдерживающие влияния на «центры сна» (II), и они блокируют восходящие активирующие влияния (III), не затрагивая возбуждений по лемнисковым путям (IV)
В качестве иллюстрации этой гипотезы рассматривается сон новорождённого — поскольку в коре ещё не имеется достаточно зрелых элементов для активного нисходящего тормозного влияния на уже значительно созревшие гипоталамические структуры, основным состоянием новорождённого является состояние сна. Причиной пробуждения является голод — возбуждаемые «голодной» кровью гипоталамические структуры реципрокно затормаживают «центры сна», устраняя их угнетающее влияние на активирующие системы, и ребёнок просыпается. После насыщения восстанавливаются прежние соотношения, и ребёнок снова засыпает.
В настоящее время после открытия целого ряда активирующих и синхронизирующих структур мозга, а также многочисленных пептидов и нейротрансмиттеров (см. далее), участвующих в регуляции цикла «сон–бодрствование», эта схема наполняется новым содержанием.











